Stejně tak to je stále poněkud záhadný tvor s některými dosud uspokojivě neobjasněnými vlastnostmi. Podávám zde proto přehled současných pohledů na tohoto živočicha založených na znalostech dokumentovaných v literatuře. Včelomorka byla poprvé zmíněna ve vědecké literatuře v roce 1740 a dodnes o ní bylo publikováno poměrně málo prací, především starých nyní už desítky let. Dále v článku uvádím výsledky vlastních výzkumů, získaných především s pomocí rastrovacího elektronového mikroskopu.
Rozšíření a morfologie
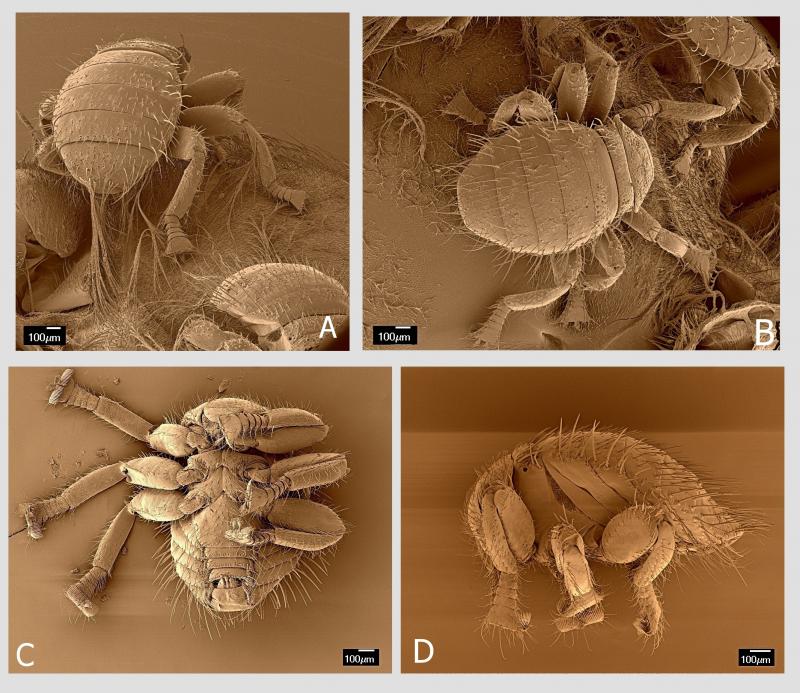
Včelomorka obecná není schopná samostatného života mimo svého hostitele. Šíří se jako nežádoucí pasažér při zalétání včel do cizích úlů, respektive hnízd cizích včelstev (Wikipedia). Dospělé včelomorky o velikosti 1–1,5 mm žijí na těle dospělých včel, především matky a dělnic, méně trubců, obvykle v počtu jednoho až několika jedinců na jednu včelu.
Vykazují extenzivní globální rozšíření: jsou dokumentovány v Africe, Asii, Evropě, Austrálii (Tasmánii), Severní a Jižní Americe (Smith and Carron 1985). Braula je velikostně i barevně podobná kleštíku (Varroa), za kterého může být omylem zaměněna. Na rozdíl od něj je mobilnější a má jen tři páry končetin. Její tělo je vypouklejší a velmi mírně protáhlé.
Imaga včelomorek pokrývají výrazné ostnité štětinky. Samečci jsou nepatrně menší než samičky. Ačkoliv jsou to mouchy, nemají na hrudi první pár křídel ani haltery.
Problém očí
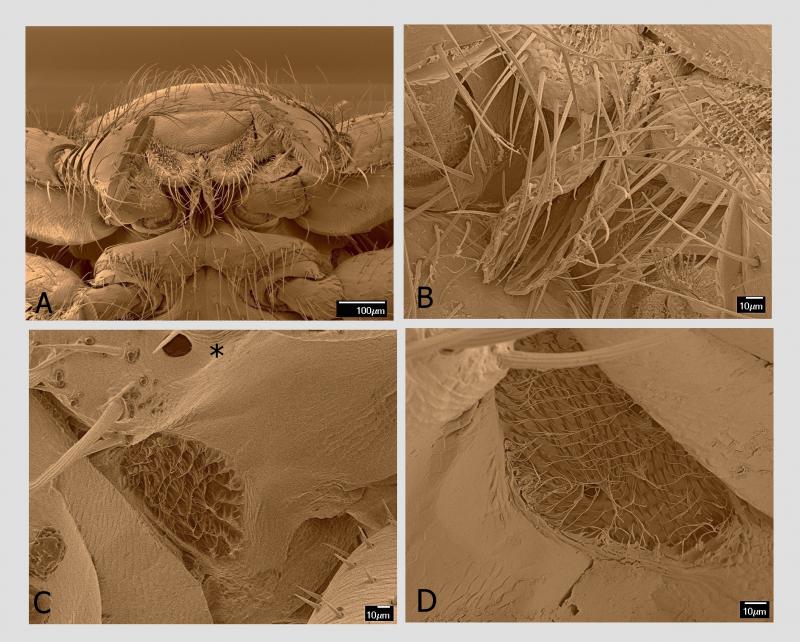
Včelomorky nemají na příčně protáhlé hlavě oči. Výrazné dvě vyvýšeniny po stranách hlavy sice oči připomínají, ale ve skutečnosti jsou to báze modifikovaných tykadel. Je zvláštní, že několik autorů (Müggenburg 1892; Massonnat 1909; Philips 1925; Ellis et al. 2010; Zapata-Carvajal et al. 2016 a další) přesto tvrdí, že včelomorky rudimenty očí, tzn. složených očí, na hlavě mají. Údajně jsou nad tykadly ve formě světlých puntíků obklopených pigmentovanými chitinovými kroužky.
Müggenburg (1892) se dokonce domnívá, že dioptrický aparát je jen slabě vyvinutý a že chitin pokrývající části hlavy, kde předpokládá složené oči, je tenký a transparentní. Nenalezl však náznaky facet, tedy nenalezl ommatidia pod neúplnou rohovkou (cornea).
Phillips (1925) odhadl z řezů hlavou včelomorky, že se tady jedná o prvotní vývojové stadium hmyzího oka, kde masa hypodermálních buněk má tendenci se radiálně uspořádat. Pigment nenalezl. Zato nalezl tenký optický nerv dosahující k tomuto rudimentárnímu oku ze supra‐oesophageálního ganglia.
Massonnat (1909) popsal tyto oční struktury ještě detailněji, ale na rozdíl od předchozího autora nalezl pigment. A jakési základy očí našli i další autoři. Já jsem ale nic takového na hlavě včelomorek nezjistil a to ani s pomocí rastrovacího elektronového mikroskopu. A nevšimli si toho ani Dietz et al. (1971), kteří jako první podrobněji studovali včelomorku právě v tomto typu elektronového mikroskopu. Přítomnost očí vylučuje také Procházka (1930). Vzhledem k tomu, že zatím ani neexistuje fyziologická práce prokazující, že včelomorky reagují na světelné stimuly, zůstává otázka přítomnosti základů složených očí nadále otevřená. Ocelli rovněž nejsou vyvinuty.
Tykadla
Aristátní tykadla jsou zasazena po straně hlavy v hlubokých prohlubních (fovea). Sestávají z velkého bazálního tykadlového článku (scapus), který je v tykadlové prohlubni blíž k centru hlavy, na nějž navazuje mohutný hruškovitý druhý tykadlový článek (pedicellus), který je v tykadlové prohlubni blíž k okraji hlavy (délka této prohlubně je 180 µm). V něm sídlí smyslový Johnstonův orgán, který pravděpodobně hraje u včelomorek důležitou úlohu v registraci akcí v okolí. Tenká, protáhlá arista (150 µm dlouhá), sestávající z několika článků, navazuje na pedicellus. Všechny tykadlové články jsou pokryty smyslovými štětinkami několika typů.
Ústní orgány
Ústní orgány včelomorek se nacházejí na přední části hlavy pod labrem a jsou redukovány do trubicovitého proboscis o délce cca 80 µm a průměru 40–50 µm a dvou skleritů v jeho okolí. Proboscis je na vnější straně částečně otevřený. Na vnitřní straně má nevýrazné záhyby. Na jeho vnější straně jsou smyslové štětinky (patrně chemosenzory). Na straně vnitřní je rovněž několik smyslových štětinek.
Hruď a končetiny
Hruď včelomorek je krátká na dorzální (hřbetní) straně, kde se neukazují všechny sklerity. Ellis et al. (2010) popisují, že zde chybí scutellum. Na hrudi jsou ve srovnání s jinými druhy hmyzu tři páry relativně mohutných, stejně velkých a podobně tvarovaných končetin, které u včelomorek představují zásadní adaptaci umožňující jim bezpečný pohyb po včele.
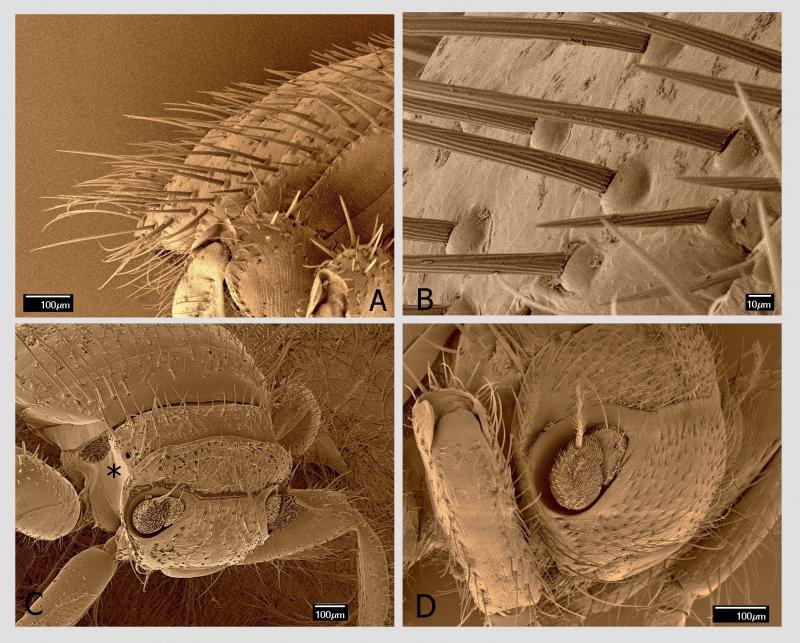
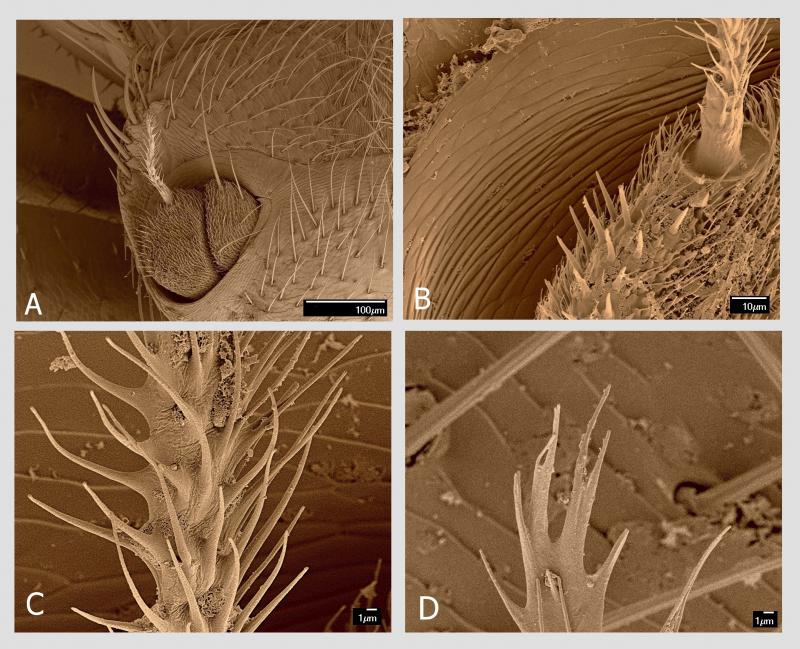
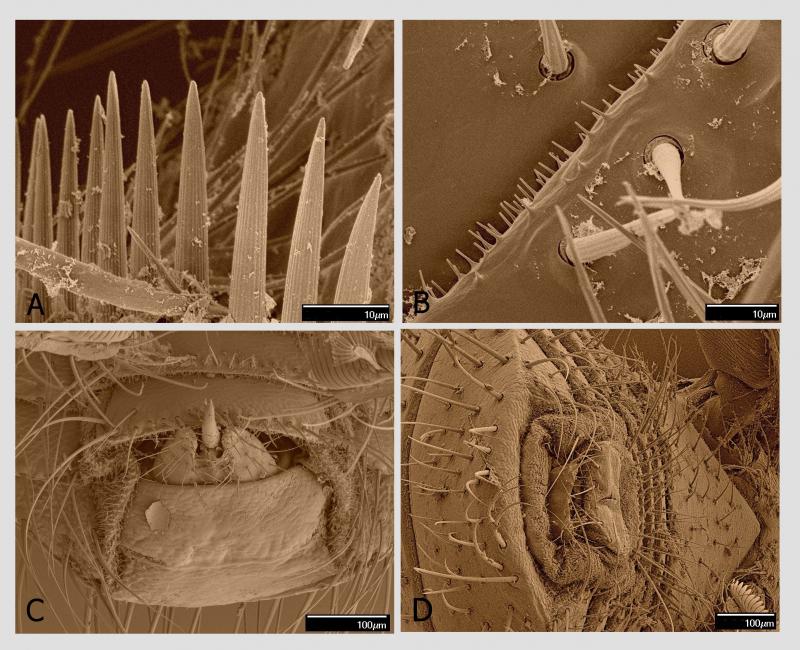
Tou specifickou adaptací jsou chodidla (tarzy). Ta jsou u včelomorek složena z pěti článků. Poslední chodidlový článek, pretarsus, je nejmohutnější z chodidlových článků a má tvar trapezoidu se širší apikální stranou. Na této straně se nachází nápadně velký hřebenovitý útvar, příčně orientovaný, o délce 230 µm. Když se pozorně podíváme, vidíme, že je uprostřed rozdělen na dva útvary. Každý tento útvar představuje modifikovaný drápek (unguiculus). Každý drápek má variabilní počet plochých a podélně rýhovaných trnů. Strukturování drápků na podobné trny, avšak v menším počtu, lze zjistit i u jiných much (Petersen et al. 2018). Jejich počet bývá od 14 do 16 na jeden drápek (zde jsem ve shodě s dalšími badateli). Právě tyto hřebenovité útvary na drápcích umožňují včelomorce efektivní pohyb po včelím těle. Fungují na principu „hrabiček zanořených hluboko do trávníku až k povrchu půdy“. Travní rostliny (analogicky všudypřítomné včelí štětinky) vnikají do mezer v hřebenovité části hrabiček (analogicky mezi hřebenité drápky) a toto spojení je velmi pevné. Kdo má zahradu, ví, o čem mluvím.
Včelomorka se nezachytává drápky na rozvětvených štětinkách všudypřítomných na povrchu těla včely, ale zachytává se na jejich bázi těsně nad kutikulou tělních článků. Názorně je to vidět na obrázcích z rastrovacího elektronového mikroskopu. Tento mechanismus je velmi efektivní, to však není všechno.
Na přední části posledního článku chodidla uprostřed hřebenitých drápků vyčnívají dvě členitě utvářené pulvily. Na rozdíl od jiných skupin hmyzu, kde jsou pulvily jednoduše měchýřkovité, zde jsou strukturované s adhezivním polštářkem (adhesive pad) na spodní části tvořeným adhezivními setami.
Tyto specializované sety s kulatým až oválným diskem na konci (spatula, průměr 2–3 µm) slouží k výkonné adhezi na podklad. Využívají se zde především van der Waalsovy síly. Adhezivní sety u členovců a způsob adheze na podklad tvoří v posledních letech předmět řady publikací. Za všechny jmenujme příklad u jiné mouchy žijící ektoparazitickým způsobem života (Petersen et al. 2018). Argo (1926) i další autoři publikovali názory o podílu těchto pulvil na pohybu včelomorek na včele. Jiní autoři, např. Dietz et al. (1971) tvrdí, že pulvily nejsou schopny přispět k adhezi včelomorek na povrchu včely, a považují jejich sety za smyslové orgány. V každém případě lze očekávat, že pohyb včelomorky po povrchu včely, byť velmi efektivní, je také energeticky náročný.
Zadeček a pohlavní orgány
Zadeček mají včelomorky oválný a kompaktní. Obě pohlaví se od sebe liší počtem viditelných abdominálních sternitů.
Samci mají čtyři ventrální abdominální sternity. Samčí vnější pohlavní orgán, aedeagus, vyčnívá z konce zadečku.
Samice mají na rozdíl od samců pět ventrálních abdominálních sternitů. Cerky jsou širší než delší, přičemž mělce vykrojený apikální okraj není pigmentovaný (Zapata-Carvajal D. et al. 2016). Na konci zadečku samic v době, kdy se v něm tvoří vajíčka, je možné pozorovat transparentní vychlípeninu, v níž lze při podrobném zkoumání nalézt rudimenty dalších tří segmentů, které jsou ale v klidovém období zanořeny do těla (Phillips 1925).
Vnější pohlavní orgány jsou u obou pohlaví na konci zadečku (ventrálně). Penis (aedeagus) samců o délce přes 50 µm vyčnívá z vykrojených pohlavních přívěsků. Cerky samic jsou rovněž vykrojené a zakrývají další části vnějších genitálií.
Smyslové orgány
Tělo včelomorky je na smyslové orgány bohaté tak, jak se na slepého epibionta sluší. Celé tělo pokrývají štětinky, na zadečku výrazně ostnité, z nichž některé dosahují délky až 300 µm. Na hlavě jsou smyslové štětinky kratší.
Tělní senzily patří do několika typů. Převážně jde zřejmě o mechanoreceptory, některé mají pravděpodobně další funkce (chemoreceptorické, teplotní a další).
Pak jsou tu části těla pokryté specializovanými štětinkami majícími vztah k funkci té které části těla. Jsou to především tykadla, která jsou prakticky celá pokrytá štětinkami. Kromě společných jsou zde i jiné typy, třeba na bazálních článcích tykadla na jeho části distální (arista). Význam tykadel pro smyslové vnímání je u hmyzu zásadní.
Další specializované štětinky jsou na ústních orgánech, na pohlavních orgánech, na končetinách. Na zadním okraji zadečkových sternitů jsou jednoduché řady malých (kolem 3 µm) smyslových štětinek se zatím neznámou funkcí (proprioreceptory?).
Při velkých zvětšeních, která umožňuje rastrovací elektronový mikroskop, jsem objevil specializovaný smyslový orgán na boku druhého hrudního článku mezi bází končetin druhého a třetího páru a průduchem. Samo kulaté až oválné stigma má délku 15–20 µm, přičemž na těle je jediný pár na tomto článku, pod povrchem průduchu je pak několik smyslových senzil. Specializovaný smyslový orgán je mělká jamka s nepravidelně oválným obvodem o průměru cca 100 μm hustě vyplněná jemnými smyslovými štětinkami o délce 10–15 µm s nevýraznou bází. Vzhledem k velikosti orgánu s mnoha desítkami senzil uvnitř lze předpokládat, že se jedná o významný smyslový orgán. Funkce je zatím neznámá.
Fyziologie včelomorky je zatím většinou neprozkoumaná a nabízí velkým prostor pro budoucí výzkumy.
Životní cyklus
Včelomorka je parazit specializovaný výlučně na život ve včelstvu. Dospělci kladou vajíčka na stěny polozavíčkovaných buněk medných plástů. Vylíhlé larvy se živí pylem a medem. Ve víčkách buněk si vrtají chodby a vytvářejí tak na nich světlé nitkovité kresby viditelné pouhým okem. Po zakuklení se po určité době líhne dospělec a přechází na tělo včely. Vývoj trvá asi 21 dní (Wikipedia). Kopulace včelomorek probíhá převážně na matkách (Phillips 1925).
Vajíčka včelomorek jsou bělavá, oválného tvaru s lemem po stranách. Velikost vajíček varíruje od 0,42 mm do 0,84 mm (délka) a 0,28 mm až 0,33 mm (šířka bez laterálního lemu). S lemem měří v průměru na délku 0,84 mm a na šířku 0,42 mm (Coffey 2007; Ellis et al. 2010).
Včelomorky kladou vajíčka do různých míst včetně prázdných i zavíčkovaných buněk, do úlomků vosku atd. Pouze vajíčka nakladená na zavíčkované plásty se vylíhnou a to po 2 až 7 dnech po nakladení v závislosti na teplotě.
Larvální vývoj
Larvy mají po vylíhnutí z vajíčka červovitý tvar se špičatou hlavovou částí a zploštělým koncem zadečku. Jsou velmi malé a lze je jen těžko vidět bez silnější lupy (Ellis et al. 2010).
Hned po vylíhnutí se snaží provrtat pod víčka. V zavíčkovaném medném plástu, kde proběhla reprodukce včelomorky obecné, lze pozorovat síť chodbiček a komůrek. Tady se larvy živí zásobami potravy pro včelí larvy: medem a pylovými zrny. Larvy procházejí třemi instary před zakuklením. Celková délka larválních stadií je 7–11 dnů podle teploty (Coffey 2007; Ellis et al. 2010).
Larvy včelomorek se kuklí v buňkách plástů v blízkosti včelích kukel. Kukly včelomorek mají žlutobílou barvu a jejich velikost varíruje od délky 1,4–1,7 mm a šířky 0,5–0,75 mm (Coffey 2007; Ellis et al. 2010). Kukelní stadium trvá 1 až 3 dny; na jeho konci se líhne dospělec (imago). Imaga včelomorek se líhnou před vylíhnutím imag včel, na která se pak přesunou.
Celkově vývoj z vajíčka do dospělce trvá od 10 do 23 dní v závislosti na teplotě. Dospělé včelomorky také přezimují na dospělých včelách (Coffey M. F. 2007).
Potravní specializace dospělců včelomorek
Včelomorka je specializovaný hnízdní potravní parazit (kleptoparazit), který je potravně vázaný výlučně na hnízdo včely medonosné. Votýpka a Urfus (2018) upřesňují, že se jedná o epibiontní forézii kombinovanou s kleptoparazitismem.
Dospělci včelomorky se zdržují na povrchu těla včel především v blízkosti ústního otvoru, nechávají se jimi přenášet a živí se tím, že když si včely předávají potravu a vzájemně se krmí (trofolaxe, trofalaxie), odsávají jim sladké šťávy od ústních orgánů. Nejčastěji lze včelomorky nalézt na včelí matce, poněvadž matka je krmena sekretem na živiny nejvydatnějším. Phillips (1925) podrobně dokumentuje způsob, jakým získává včelomorka potravu od včely. Včelomorky se zdržují na včele především na hřbetní straně zadečku a hrudi, dokážou se ale rychle přesunout na hlavu k ústním orgánům (Coffey 2007). Pohyb po včele není snadný a včelomorky proto mají velmi specifické morfologické adaptace na končetinách, které jim takový pohyb umožňují.
Existují důkazy, že včelomorka dokáže u včely indukovat regurgitaci (návrat polknuté potravy zpět do úst) stimulací horní části včelího labra, až včela vysune sosák. V tom momentě včelomorka saje potravu a další sekrety, které takto včela víceméně nechtěně poskytne (Ellis et al. 2010). Další autoři popisují podobné mechanismy, jak včelomorka stimuluje včelu, aby od ní získala trochu potravy. Tyto procesy jsou ale velmi rychlé, a tak zatím dosud chybí přesné údaje. Svoji roli zde může hrát stimulace včelích tykadel pomocí pulvily na končetinách včelomorek (Dietz et al. 1971). Včela pootevře kusadla a dovolí včelomorce zasunout trubicovitý proboscis do ústní dutiny, kde najde potravu.
Sosák včelomorky je jednoduchý a má tvar částečně otevřené trubice. Byl podrobně prostudován (například Massonnat 1909) a bylo zjištěno, že není schopen proděravět kutikulu tělních segmentů včely, a to ani tenčí intersegmentální membránu. Chybí zde tuhé stylety na proboscis. Včelomorka se proto nemůže živit hemolymfou včel, anebo tukovým tělesem jako kleštík.
Ve svých experimentech jsem zjistil, že včelomorky přítomné u skupiny včel umístěných delší dobu v transportních klíckách opustily včely v době, kdy došla zásobní potrava (medocukrové těsto) a skoro všechny uhynuly v těsném okolí klícek. Jen některým se podařilo dostat se dál (několik centimetrů) od klícek, uhynuly pak rovněž. Procházka (1930) uvádí, že mladé včelomorky, které opustily včelu, žijí ještě čtyři dny. Staří jedinci hynou záhy. Lze proto konstatovat, že včelomorka je se včelou úzce potravně svázaná.
V minulosti byl zhotoven roztěr zažívací soustavy imag včelomorky (Phillips 1925). Autor zjistil v jednom jedinci 11 pylových zrn. U včel našel ve stejné době velké množství pylových zrn. Pyly u obou druhů byly stejného druhu. Dosud ale není známo, zda pyly jsou důležitou součástí potravy včelomorek.
Rozsah napadení včelomorkou
Pokud se týká počtu včelomorek, které mohou být přítomny na jedné včele, tak z vlastních pozorování vím, že jde obvykle o jednoho až několik jedinců. Někdy jde o větší počet včelomorek, zvláště u matek (až 40 jedinců). Weems (1983) však popsal případ, kdy na jedné matce bylo 187 včelomorek; podle jiných autorů mohlo jít o jiné parazity, především kleštíky. Souhlasím s autory Dietzem et al. (1971), že je třeba tuto otázku dále studovat.
Míra škodlivosti včelomorky pro včely
Včelomorka obecná, jak vyplývá z výše uvedeného, nepatří mezi nebezpečné parazity. Na zpomalování rozvoje včelstva, eventuálně na jeho úhynu ve skutečně extrémním případě nebo uhynutí matky by se mohla včelomorka podílet jen při extrémním přemnožení v oslabeném včelstvu. Velký počet včelomorek přítomných na těle včely totiž může včelu rušit. U matky může snižovat množství přijímané potravy a jejím rušením snížit počet kladených vajíček (Coffey 2007).
Protivníci včelomorky
Od konce 20. století, kdy se aplikují syntetické jedy proti kleštíku včelímu, těchto drobných bezkřídlých much citelně ubylo. Nesprávně bývá výskyt včelomorek u včel označován za chorobu, tzv. braulózu, na což upozornil Přidal (2007).
Včelomorka má jednoho predátora, jímž je štírek knihový (Procházka 1930).
Poděkování
Poděkování patří Mgr. Bronislavu Grunovi, který zásadně přispěl ke vzniku tohoto článku, protože autorovi zaslal několik desítek živých včelomorek ke studiu, a Ing. Petru Texlovi za poskytnutí některých fotografií.
Literatura
1. Argo V. N. 1926. Braula coeca, or Bee Louse, Gl. Bee Cult. 54: 435–438.
2. Coffey M. F. 2007. Parasites of the Honeybee. Department of Agriculture, Fisheries and Food, 77 str.
3. Dietz A., Humphreys W. J., Lindner J. W. 1971. Examination of the bee louse, Braula coeca, with the scanning electron microscope. Apiacta 1.
4. Dobson J. 1999. A bee-louse Braula schmitzi Orosi-Pal (Diptera: Braulidae) new to the British Isles, ans status of Braula spp. in England and Wales. Ent. Nat. Hist, 139–147.
5. Ellis J. D., Mortensen A. N., Zettel Nalen C. M. 2010. Bee Louse, Bee Fly, Braulid, Braula coeca Nitzsch (Insecta: Diptera: Braulidae). EENY 472, University of Florida, IFAS Extension, http://entomology.ifas.ufl.edu/creatures.
6. Massonnat S. 1909. Contribution a L'etude des Pupipares. Ann. de L'Univ. de Lyon, vol. 1, Sciences, medicine, fasc. 28: 1–388.
7. Morse R. A., Flottum K. (eds.) 1998. Honey Bee Pests, Predators, and Diseases. 3rd edition. Medina, OH: A. I. Root Company.
8. Müggenburg F. H. 1892. Der Rüssel der Diptera pupipara. Archiv für Naturgeschichte, 58 (1): 287–332.
9. Petersen D. S., Kreuter N., Heepe L., Büsse S., Wellbrock A. H. J., Witte K., Gorb S. N., 2018. Holding tight to feathers – structural specializations and attachment properties of the avian ectoparasite Crataerina pallida (Diptera, Hippoboscidae). J. Exp. Biol., (2018) 221: 1–9.
10. Peterka V. 1930. O včelomorce. Český včelař 64: 14–19.
11. Phillips E. F. 1925. The bee-louse, Braula coeca, in the United States. USDA, Department circular 334, 1–12.
12. Přidal A. 2007. Parazitismus, nemoci včel a názvosloví živočichů. Moderní včelař, 1: 27–29.
13. Smith I. B. Jr., Caron D. M. 1985. Distribution of the beelouse, Braula coeca, in Maryland and worldwide. American Bee Journal 125: 294–96.
14. Voigt D, Tsuchihara K., Yazaki K., Kuroda K., Tsubaki R., Hosoda N. 2017. Strongest grip on the rod: tarsal morphology and attachment of Japanese pine sawyer beetles. Zool Lett. 2017; 3: article no. 16.
15. Votýpka J., Urfus T. 2018. Epibionti aneb Život na životě. Živa 4 (2018):85–88.
16. Weems H. V. Jr. 1983. Beelouse, Braula coeca Nitzsch (Diptera: Braulidae). Entomology, Circular No. 252, July 1983.
Zapata-Carvajal D., Chérrez-Neacato E., Chávez-Larrea M., Saegerman C., Ron-Román J. 2016. First report of the bee louse – Braula coeca (Diptera: Braulidae) in apiaries of Los Chillos Valley, Province of Pichincha, Ecuador. International Congress of Entomology, Orlando, Florida, USA, September 25–30.